 Chapter 7
Chapter 7 
VISION
Photoreception is a particularly important sense for most primates, including man, but it is not unique to primates or even mammals. Even mollusks have photoreceptors, but one may question whether they possess vision in the same sense as we have it. Most objects reflect light, and because light travels at high speed, it is possible to nearly instantly assess their shape, size, position, speed, and direction of movement. The light rays emanating from an object are gathered and focused onto an array of photoreceptors. Activities generated in the different photoreceptors by the light interact to produce a two-dimensional representation of the object which is transmitted to the brain. The brain then reconstructs a three-dimensional representation using information received from the two eyes. The end-products of the activity of the visual system are sensations that represent the object and its surroundings. These sensations can be used to guide our immediate behavior, or they can be stored for future reference. Visual sensations contain a great deal of information, and understanding these complex phenomena is no simple matter. The best place to begin the study of vision is at the eye itself.
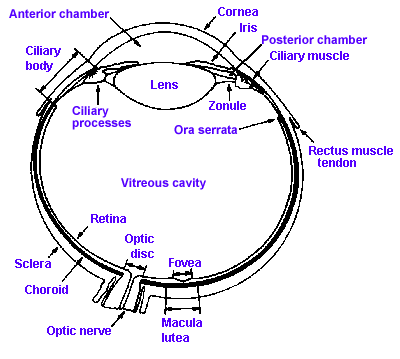 |
Fig. 7-1. A section through the human eye illustrating the major structures. (Walls GL: The Vertebrate Eye and its Adaptive Radiations. New York, Hafner, 1967) |
Figure 7-1 shows a cross section through the human eye. It consists of two fluid-filled chambers separated by a transparent structure, the lens. Nearly the entire eye is covered with a tough, fibrous coating called the sclera that is modified anteriorly to form the transparent cornea. The human cornea is about 12 mm in diameter, about 0.5 mm thick in the center and 0.75 to 1 mm thick on the edge, and it is made of the same collagenous connective tissue substance as is the sclera, but the fibers of the cornea are oriented in parallel arrays that let light pass through with minimal scatter, whereas fibers of the sclera are random and light rays are scattered when passing through. The result is that light passes easily through the cornea, but not through the sclera. Lining the inside of the posterior two-thirds of the sclera are two membranes: the choroid, a pigment layer containing the vascular supply for the eyeball as well as mechanisms for maintaining the integrity of the photoreceptors, and the retina that contains the photoreceptors and other neural elements essential to our visual process. The fine structure of the retina will be considered in detail later.
The human lens is about 11 mm in diameter and 3.5 mm thick at its thickest point, and it is suspended in place by zonule fibers that attach to the ciliary process anterior to the retina. A set of smooth muscle fibers, the ciliary muscle, lie between the ciliary process and sclera. Just anterior to the lens is a pigmented structure called the iris, that is like the diaphragm on some cameras in that it has a hole in the center of variable aperture, the pupil. The pupil is surrounded by two sets of muscles, one that encircles the aperture, the sphincter pupillae, and one that runs radially out from it, the dilator pupillae.
The anterior chamber of the eye is filled with aqueous humor, a watery fluid of low protein content that is formed from plasma. The vitreous cavity contains a gelatinous substance, the vitreous or vitreous humor. In many people the vitreous is not completely clear, but contains particulate matter that is not transparent. This material may be stationery or may float around, producing "spots before the eyes," the floating variety being called "floaters."
At this point, we should spend just a moment reviewing the properties of light, which is a form of electromagnetic radiation. Light is radiant energy emitted by matter when it is at high temperature, when it is electrically excited, and when it is undergoing certain chemical or physical processes. Early on, physicists attempted to account for the properties of light as a stream of particles emitted by a luminous object. Later, they attempted to account for them in terms of waves, but light has properties both of waves and of particles(1). In any case, light behaves as if it has characteristic wavelengths that are measurable. The total spectrum of wavelengths of electromagnetic radiation is quite broad, ranging from 1 nanometer (10-9 m, abbreviated nm) to over 104 m, yet we see only a small part of that spectrum (400-700 nm, Fig. 7-2). Some other animals have a broader sensitivity than man, being able to see in the ultraviolet region (less than 400 nm), but not even these animals see the whole light spectrum, 380-760 nm.
 |
Fig. 7-2. The spectrum of light visible to the human eye. |
Light travels in straight lines from its source unless it passes through a high intensity electro-magnetic field or from a medium of one refractive index into a medium of another. In the first case, the light is bent along a curved path. We will not consider this further because it plays no role in the majority of normal human vision. When light passes from one medium into another of a different refractive index, the direction of travel of the light beam is changed at the interface. This is the basis for the phenomenon of refraction, that we will discuss in detail in the next section.
Psychologically, light has three aspects we need to consider. It has hue, a quality correlated with wavelength and analogous with pitch in hearing. It has saturation, a quality related to homogeneity or purity of wavelength and comparable to timbre or freedom from noise in audition. The color of a light is a function both of its hue and saturation. Finally it has brightness, a quantitative measure of the intensity of the light and somewhat analogous to loudness in hearing. The intensity of light is dependent upon the amplitude of its waves or alternatively the number of photons (particles of light).
| Optical Functions of the Eye |
The process of light reception and transmission has a number of parts. First, we will consider the optical functions of the eye: accommodation(2), the process of forming a focused image, and control of the amount of luminous flux at the retina.
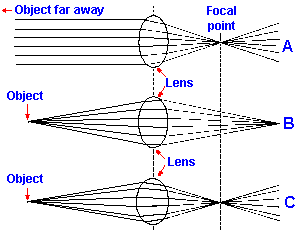 |
Fig. 7-3. Convergence of light by a convex lens. A. Light from an object far away is brought to focus at the focal point of the lens. B. Light from a nearer object is brought to focus at a point farther way than the focal point of the lens. C. The radius of curvature of the lens must be decreased to bring the light to focus at the focal point of the lens in A. |
Accommodation
A convex lens like that in the eye has the property of bending penetrating rays of light toward its axis (Fig. 7-3A). This property results from the fact that light travels more slowly in some materials, e.g., water and glass, than in air (and more slowly in air than in a vacuum). The amount of slowing in a material is specified by its refractive index, which is the ratio of speed of light in a vacuum (the speed of light in air is slightly less, but very close) to the speed of light in the material (refractive index = speed in vacuum/speed in material). If light passes from one medium to another at an angle to the interface between them, the rays are bent as shown in Figure 7-3A, where parallel rays of light emanating from an object far away are bent as they pass through the convex lens in such a way that they converge onto a point in space. Where they converge depends upon the strength of the lens which is specified by its focal length. The focal length of a lens is the distance from the center of the lens to the point of focus of light from a distant object. Focal length is a function of the refractive index of the lens material and the radius of curvature of the lens. The larger the radius of curvature, i.e., the less spherical the lens, the longer the focal length is.A convenient way to specify the focal length or strength of a lens is in terms of the diopter, the reciprocal of the focal length of the lens expressed in meters. As you know, there are two kinds of lenses, convex lenses (converging lenses), like that shown in Figure 7-3, and concave lenses (diverging lenses). With a convex lens the image of an object is formed on the side of the lens opposite from the object. With a concave lens no actual image is formed, but physicists identify a virtual image that lies on the same side of the lens as the object. Because the images lie on opposite sides of the lenses in the two cases, convex lenses are assigned positive diopter values (presumably for their real images) and concave lenses are assigned negative diopter values. For example, a convex lens with a focal length of 0.5 meter is a +2 diopter lens, whereas a concave lens with a focal length of 0.33 meter is a -3 diopter lens. The higher the absolute value of the diopter rating of a lens, the stronger it is, i.e., the more it converges or diverges light that enters it.
When objects are close, the distance from the lens to the point of focus varies with the distance to the object. Thus, the light from a source farther away in Figure 7-3A is brought to focus closer to the lens than that from a source nearer the lens in Figure 7-3B. In the eye, the distance from the lens to the retina is fixed so that in order to focus the light from a near source and also light from a far source the strength of the lens must be increased (i.e., the lens made more spherical), a process called accommodation(3) and illustrated in Figure 7-3C. The lens is a flexible structure that, when released from the tension of the zonule, becomes nearly spherical in shape. Normally the lens is under considerable tension and thus is flattened, a condition appropriate for focusing objects at a distance. Under these conditions, light from near objects is brought to focus behind the retina but is unfocused on the retina, and the images of them are blurred. Because of the way in which the ciliary muscle is attached (see Fig. 7-4), its contraction results in a relaxation of the tension on the lens, causing the lens to become more curved (i.e., more spherical), thereby bringing nearer objects into focus on the retina. By grading the amount of contraction of the ciliary muscle and therefore the tension on the lens, the nervous system can focus objects located from about 9 cm from the eye to infinity. Images closer than 9 cm cannot be focused by the human eye.
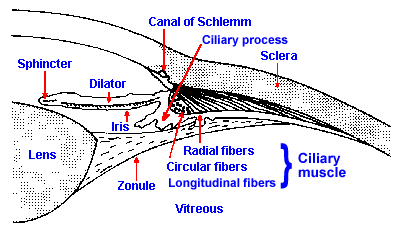 |
Fig. 7-4. The anterior portion of the eye. The arrangement of the lens, the zonule, and the ciliary muscles is illustrated along with the position of the iris and the arrangement of the muscles asociated with it. (Weymouth FW: The eye as an optical instrument. In Ruch TC, Patton HE [ed]: Physiology and Biophysics, 19th ed. Philadelphia, WB Saunders, 1965) |
Control of the ciliary muscle is exercised by the autonomic nervous system which uses information obtained from the retina itself to adjust the focus of an image. Presumably, the pathway for producing accommodation must involve the retina, optic nerve, lateral geniculate nucleus, visual cortex, superior colliculus(4) and finally the oculomotor nucleus, relaying in the ciliary ganglion to reach the ciliary muscle. Participation of visual cortex is inferred from the fact that accommodation is normally accompanied by pupillary constriction (miosis of accommodation) and convergent eye movements, in conjunction with the observation that stimulation of visual cortex results in pupillary constriction. The nerve fibers supplying the ciliary muscles are parasympathetic postganglionic fibers and cholinergic (they use acetylcholine as a transmitter substance). Transmission through the cholinergic junction is readily blocked by atropine, which produces relaxation of the ciliary muscles so the lens is flattened and focused for distant objects. Ophthalmologists frequently relax the ciliary muscles to test the eye for refractive errors, but they use a shorter acting parasympathetic blocking agent, such as tropicamide. They do this in order to put the eye in a standard condition for comparison with a standard normal eye.
Actually, the lens is not strong enough by itself to focus a parallel beam of light on the retina. The cornea also acts as a lens (of fixed strength) and accounts for about 75% of the refractive strength of the optical system. The total refractive power of the eye is about +60 diopters. The cornea accounts for about +45 diopters, the lens only +17 diopters, and the remainder of the +60 is accounted for by the liquid-tissue interfaces. (Note: Some of the interfaces are from regions of higher refractive index, e.g., cornea 1.376, to regions of lower refractive index, e.g., aqueous humor 1.336, and thus act as diverging lenses, in this case with a power of -5.9 diopters.) The lens has a higher refractive index than the cornea (1.386 versus 1.376), but it is surrounded by fluids with refractive indices much higher than that of the air (1.000) at the anterior boundary of the cornea. It is the ratio of the refractive indices at an interface that determines the refractive power of that interface. The greatest ratio, and therefore the greatest refraction, occurs at the air-cornea interface. With accommodation, the eye gains another +10 to +12 diopters depending upon the person's age. Removal of the cornea requires a strong convex lens (about +45 diopters) to correct for the loss in refractive power, but a weak man-made lens will correct for loss of natural lens function. The real problem in loss of lens function is the inability to refocus from near vision to far vision (i.e., to accommodate), a fine adjustment that the lens provides.
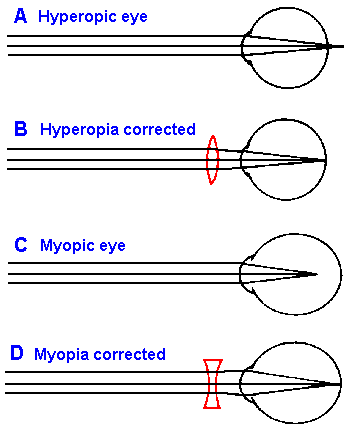 |
Fig. 7-5. Errors of refraction. A. The hyperopic eye with convergence of parallel light behind the retina. B. The hyperopic eye corrected with a convex lens. C. The myopic eye with convergence of parallel light in front of the retina. D. The myopic eye corrected with a concave lens. |
Errors of refraction
With increasing age, the lens loses some of its elasticity so that even though the ciliary muscles contract and release tension on the zonule, the lens cannot round-up as it should. The result is that near objects cannot be properly focused, calling to mind the image of a man in his 40's or 50's (the age at which this condition becomes apparent) holding a book at arms length in order to read the small print. The nearest point at which the eye can distinctly perceive an object is referred to as the near point. At 10 to 30 years of age, the near point is about 9 cm in front of the eye. It gradually recedes from the eye, being about 28 cm at 45 years and 80 cm at 60 years. There is a gradual loss in ability to see close objects. This condition is called presbyopia or old-sightedness. Presbyopia is related to a general condition called hyperopia or far-sightedness in which images are formed behind the retina. Hyperopia, shown in Figure 7-5A, results when the length of eye is too short for the refractive power of the lens; this can result from having too short an eyeball or too weak a lens system. When a weak lens system results from age-related changes in the lens itself, the condition is presbyopia. The hyperopic eye sees distant objects clearly only by using the normal accommodative mechanism of the lens. It does this at the expense of constant contraction of the ciliary muscles, which produces eye strain. Further, because part of the accommodative process is used to see even distant objects, less is available for viewing near objects; very near ones are never in focus.Because the image is focused behind the retina in hyperopia, the correction is accomplished by increasing the strength of the lens system, converging the light beam even more. This involves putting another convex (or positive, in terms of diopters) lens in front of the eye, as shown in Figure 7-5B.
Another common deviation from the emmetropic, or normal eye, occurs when the image comes to focus in front of the retina as illustrated in Figure 7-5C. This condition, called myopia or near-sightedness (because near objects are readily focused), results when the eyeball elongates too much for the refractive power of the lens, most commonly, myopia occurs beginning at puberty. For the myopic eye there are always points sufficiently near, from which light diverges enough to be brought to focus upon the retina, but light from all points at greater distances cannot be focused. Ophthalmologists use the term far point to refer to the remotest point at which an object is clearly seen by the unaccommodated eye. In the myopic person, the far point is closer to the eye than normal. The optical correction for this condition is shown in Figure 7-5D. A concave (or negative, in terms of diopters) lens of the appropriate power (refractive index and curvature) will diverge light rays from more distant objects enough to compensate for the over-convergence of the eye and bring a focused image onto the retina of the lengthened eyeball.
Three other corrections for myopia have become popular. Radial keratotomy, making radial cuts in the cornea to flatten it out, can correct about 3 diopters of excess power, but the procedure has several disadvantages. (1) It is difficult to accurately control the depth and length of the cuts, making the results somewhat unpredictable. In addition, too deep a cut can damage the endothelial layer on the inside of the cornea, the function of which is to pump water out of the cornea. This layer is essential to normal corneal function; humans cannot regenerate it. (2) Epikeratophakos involves using the cornea from a donor eye as a living contact lens. The donor cornea is sewn to the recipient cornea. Some success has been achieved with this procedure. (3) The third procedure involves fitting a special contact lens to the cornea. This lens "reshapes" the corneal surface to change its refractive power. Considerable success has been claimed for this procedure, but it is probably too new to know the permanence of the correction.
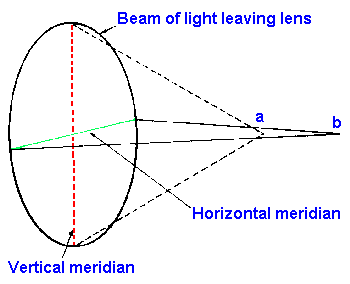 |
Fig. 7-6. The mechanism of production of astimatism. Illustrated at left is a beam of light leaving the lens. Because the radius of curvature is greater along the horizontal meridian, the light passing through that meridian is focused at point b whereas light passing through the vertical meridian of lesser radius of curvature comes to focus nearer, at point a. |
Yet another error of refraction, astigmatism results from asymmetry of the cornea (occasionally also the lens). The ideal refractive surface is symmetrical in all meridians such that a line of light (the image of a luminous line in space) would come to focus at the same distance from the lens no matter how it was oriented. When this is not the case, an astigmatism results. Figure 7-6 shows that in the astigmatic eye a beam of light emerging from the lens along the vertical meridian (the axis of least radius of curvature in our example) is brought to focus at a and along the horizontal meridian (the axis of greatest radius of curvature) at point b. If point a lies on the retina, then a line in the vertical meridian would appear focused; a line in the horizontal meridian would appear blurred. If point b were on the retina, the situation would be reversed. If the retina lies between a and b, then both would appear blurred. In the normal eye, points a and b would be the same. Astigmatism usually results from a cornea that is not spherical, but more egg-shaped with the long axis not parallel to the direction light is traveling to the retina. Correction for astigmatism is accomplished by using a combination of spherical and cylindrical lenses, usually ground from a single piece of glass in such a way that (in Fig. 7-6) refraction would be increased in the horizontal or decreased in the vertical meridian or both depending upon the location of the retina with respect to points a and b. An optometrist must determine how the asymmetry is oriented as well as how strong to make the corrective lens.
The pupil
The functions of the pupil are to produce sharp images, especially in close work; to alter the depth of field; and to help regulate the amount of light striking the retina. When the pupil constricts, it prevents light from entering the periphery of the lens. It is on the periphery where spherical aberrations, that is, unequal refraction of light by different parts of the lens, and chromatic aberrations, unequal refraction of light of different wavelengths, are greatest. Spherical aberrations occur because light passing through the edge of a lens is actually refracted more than light passing through its axis and comes to focus in front of the axial focus. The image formed on the retina of a spot of light in the visual field is a spot surrounded by a blurred halo instead of a single spot. An entire object would appear blurred under these conditions. The pupil partially reduces the influence of this aberration when it constricts, allowing light to enter only through the center of the lens.Light of long wavelengths, like red light, is refracted less than light of shorter wavelengths, like violet light, so the lens tends to separate wavelengths or colors, producing a rainbow effect(5). This effect (chromatic aberration) would be the worst on the periphery of the lens where spherical aberration is greatest. Exclusion of the periphery of the lens is important in partially reducing the formation of colored rings around an image. Constriction of the pupil or miosis also helps to reduce the rings around an image that result from light entering the eye from objects too far or too near to the lens to be in focus at a given radius of curvature.
Controlling the depth of field, that is, the range of distances over which images are in focus for a given strength of the lens, is also a function of pupil diameter. When the eye is focused on objects at a great distance, the depth of field is normally large and pupil size is relatively unimportant. When focused for near objects, however, the depth of field is small and can be increased markedly by closing down the pupil, just as it is by decreasing the aperture in a single lens reflex camera.
The pupil also changes diameter with changes in light intensity at the retina, an increase causing the pupil to constrict and a decrease causing the pupil to dilate. This response is termed, the light reflex. Within limits, the light reflex provides an adequate amount of light on the retina, increasing the amount in dim light, decreasing it in bright light. The aperture of the pupil is not a constant function of light intensity because the sensitivity of the retina changes as a result of adaptation, which we will consider shortly. Thus, when the light intensity increases moderately, the pupil constricts, but then under constant illumination it gradually dilates again. With larger changes in intensity, the pupil will remain constricted. It is not unusual for the eye to experience a million-fold change in light intensity, but the pupil is only capable of about a four-fold change in diameter(6), that corresponds to about a 16-fold change in light intensity. This is not much compared to the total range the eye can experience; the greater part of the compensation is accomplished through retinal adaptation. However, the pupillary reflex system is very fast compared to retinal adaptation that takes minutes.
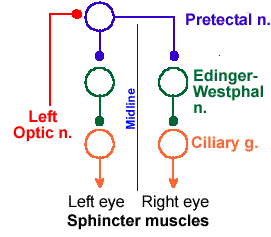 |
Fig. 7-7. The circuitry for production of both the direct and consensual light reflexes. |
The light reflex is mediated by the autonomic nervous system (Fig. 7-7). The afferent limb of the reflex involves the retina and the optic nerve. Fibers in the optic nerve give off collaterals to the pretectal nucleus, a structure just rostroventral to the superior colliculus. From this nucleus, fibers run bilaterally to the Edinger-Westphal nucleus, part of the oculomotor nucleus (third cranial nerve), forming connections with fibers of the third cranial nerve that are parasympathetic. These fibers relay again in the ciliary ganglion to innervate the sphincter pupillae. Increasing the amount of light at the retina increases the discharge to the various relays, finally causing contraction of the sphincter, closing down the pupil. At the same time the sympathetic fibers that innervate the dilator pupillae are inhibited by this increased activity, relaxing this antagonistic muscle. Decreasing the amount of light at the retina causes an increase in sympathetic activity and dilation, while it reduces the output to the sphincter through the ciliary ganglion. The pretectal nucleus connects to the Edinger-Westphal nuclei bilaterally, which accounts for the fact that both pupils normally constrict together even though the light is directed into only one eye. The constriction of the pupil through which the light is shown is called the direct light reflex, whereas constriction of the contralateral pupil is called the consensual light reflex. The existence of the consensual reflex is a great aid in determining the location of pathology involving this system. A lesion of the afferent limb of the reflex (the part conducting toward the brain, including the optic nerve up to the pretectal nucleus) results in abolition of both the consensual and direct responses evoked by directing a light in the eye on the same side as the lesion, whereas a lesion of the efferent limb (the part conducting away from the brain) results in abolition of the direct response evoked from the affected side but not the consensual reflex evoked from that side (though the consensual reflex evoked from the unaffected side will also be abolished).
The mechanism of accommodative miosis (pupillary constriction accompanying accommodation) is not identical with miosis of the light reflex, as indicated by their dissociation in certain disease processes. An example is the Argyll Robertson pupil that occurs in most syphilitic infections of the central nervous system. The pupil is constricted and is not responsive to light, but does respond normally during accommodation. The only regular pathological concomitant of syphilitic infections of the central nervous system is gliosis surrounding the aqueduct of Sylvius that has been interpreted to indicate damage to the Edinger-Westphal nucleus. This implies that there are different cells for the two kinds of miosis, one type resistant to the spirochete. However, the cause of the Argyll Robertson pupil is not certain. In diphtheritic neuritis, the converse occurs: the pupil responds normally to light, but does not show accommodative miosis.
Dilation of the pupil is a normal concomitant of emotional states such as fright or alarm. The dilator reflex is also elicited by most forms of noxious stimulation. Use has been made of the change in pupil size that occurs with pleasurable sensations in monitoring responses of patients in many clinical situations.
| Receptive functions of the eye |
The photoreceptors of the eye are located within a thin sheet of tissue, the retina. The structure of the retina is illustrated in Figure 7-8. Two types of receptors are present in the retina, the rods and the cones, so named because of their characteristic appearance. The human retina contains about 6.5 x 106 cones and 100 x 106 rods. Both rods and cones are found in the receptor layer of the retina, that outermost layer nearest the choroid. The receptors are connected to cells in the middle layer of the retina, the bipolar cells, which are in turn connected to cells in the innermost layer, the ganglion cells. The axons of the ganglion cells are the fibers that eventually come together to form the optic nerve. Also in the retina are amacrine cells, that interconnect the bipolar cells with each other and the bipolar cells with ganglion cells, and horizontal cells, that interconnect receptor cells. These latter two cell types may play a role in sharpening images by surround inhibition, and the horizontal cells may play a role in color opponent processes of color vision (see section on visual neurons in the retina). In addition, the retina contains neuroglia and blood vessels that support the tissue. It is important to note that because of the way the retina is oriented, light entering the eye reaches the receptors only after passing through the network of blood vessels, cells, and fibers. The light finally reaches the light sensitive photopigment in the outer segments of the receptor cells (Fig. 7-8), and what is not absorbed there is absorbed by the pigmented choroid, although a small part may be reflected back to the receptors.
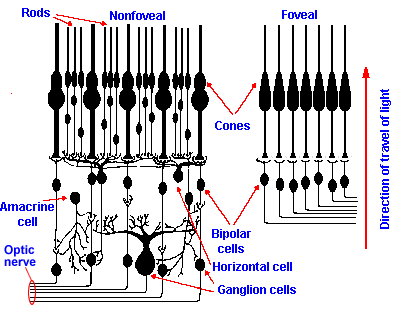 |
Fig. 7-8. The cellular organization of the foveal (right) and nonfoveal (left) retinas. Notice that higher density of cones, their one-to-one connection with bipolar cells, and the absence of ganglion and other cells in the foveal area. Rods, located only outside the fovea, converge in large numbers onto bipolar cells. Cones in the fovea are also smaller and more closely packed than in the nonfoveal retina. |
The receptors themselves consist of an inner and outer segment. The outer segment is composed of neatly folded lamellae that contain the photosensitive pigments that change their structure by isomerization when they absorb light. When they isomerize, they produce a change in the membrane potential of the receptor cell, the receptor potential. The term receptor potential is preferred here because no action potentials are generated in the receptors themselves. In fact, it is in the third-order ganglion cells that action potentials first appear in the visual system. The inner segment is joined to the outer segment by a narrow constriction that has characteristics of a true cilium. The inner segment contains mitochondria and endoplasmic reticulum and may be immediately adjoined to the cell body, as in cones, or connected to the cell body by a slender process, as in rods. The short axon arises from the soma in both rods and cones.
The retina may be partitioned into several regions that differ somewhat in structure. The central region or fovea ("pit") is a small depression of the retina (a concavity on the inner or vitreal side), where the retina is thinner than elsewhere. The fovea is about 1500 µm in diameter in man. Only cones are present in the fovea. The macula lutea or "yellow spot" is a region, including and surrounding the fovea, in which yellow pigment is found in the neuron layers of the retina. In the region surrounding the fovea, there is a gradual decrease in the density of cones and a gradual increase in the density of rods until, at the outer margin of the retina, there are almost no cones at all. Finally, there is a region in which there are no receptors, but an accumulation of the ganglion cell axons to form the optic nerve where it leaves the eye. This region, called the optic disc, forms the physiological scotoma or blind spot.
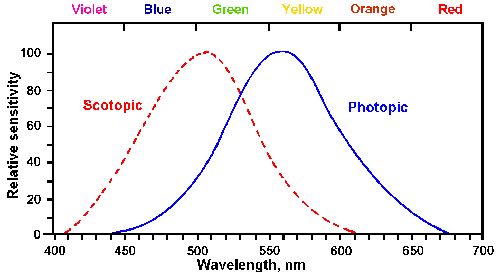 |
Fig. 7-9. Photopic and scotopic visibility curves. Threshold intensity is plotted against wavelength for the dark-adapted eye (scotopic curve) and the light-adapted eye (photopic curve). (Winton FR, Bayliss LE: Human Physiology, 5th ed. Boston, Little, Brown, 1962) |
Retinal sensitivity
Visual systems specialize in the detection of electromagnetic radiation. The human visual system is sensitive to radiation in wave lengths from 400-700 nm, that is, the spectrum of visible light from violet through red. Monochromatic sensitivity (sensitivity to one hue) varies continuously through the spectrum; thus thresholds are normally expressed as a curve called the visibility curve, relating sensitivity to wavelength. Actually, there are two such curves because the sensitivity varies with the immediate history of the eye. Figure 7-9 shows these curves. If the eye has been in darkness for at least 30 minutes, and the tests are done with brief exposures, the curve on the left results. This is called the scotopic visibility curve, and it covers the range from 400 to 625 nm with a maximum sensitivity at about 507 nm, that is, sensitivity is maximum to blue-green light. If, on the other hand, the eye has been in normal daylight, the curve to the right, the photopic visibility curve, results. The maximum sensitivity for photopic vision is at 555 nm or in yellow-green light. Notice that the values on the ordinate are normalized so that both curves reach a peak at a value of 1.0. If this were not the case, the scotopic curve would be much taller because the eye is much more sensitive after being in the dark than after being in the light. These properties are the result of the characteristics of the receptors that lie in the retina.Rods and cones not only differ in their appearance, but also in their connections to the bipolar cells and in the kind of visual pigment they contain. Some important differences are summarized in Table 7-1. It is these differences that account for many of the physiological properties of the retina. Cones in the fovea connect in a one-to-one fashion with bipolar cells that connect in a one-to-one fashion with ganglion cells, as illustrated in Figure 7-8. On the other hand, outside the fovea several rods and cones connect to a single bipolar cell, as illustrated at the left in Figure 7-8. Because many rods and cones connect to a single bipolar cell, a given amount of illumination will more strongly excite bipolar cells in the peripheral retina than in the foveal retina. In addition, the rods themselves are more sensitive to light, showing six times as much change in membrane potential per quantum of light as cones. Thus, the sensitivity of the retina is greater where there are more rods and less where there are only cones. This accounts for a phenomenon well known to sailors on watch. A very faint light seen out of the corner of the eye, where the image falls on the peripheral retina, disappears when the eyes are turned toward the light, where the image falls on cones in the fovea. The greater sensitivity of the peripheral retina gives it superiority in scotopic or night vision, which lacks color information, whereas the foveal retina normally functions in photopic or day vision, which is in color. It is not clear why there is such an abrupt shift from rod to cone dominance in the photopic condition, but it appears that rod functioning is somehow suppressed when cones are active.
Table 7-1
Comparison of Peripheral and Foveal Retina | ||
Property |
Foveal Retina |
Peripheral Retina |
| Threshold | Relatively high | Exquisitely low |
| Receptor distribution | Cones only | Rods and cones |
| Convergence | Limited or none in fovea; probably more elsewhere | Extensive |
| Illumination | Photopic (Daylight) | Scotopic (Night) |
| Functions | Central vision
Color vision Detail vision |
Peripheral vision
Achromatic vision Poor detail vision |
Acuity
The measure of acuity or detail vision is the minimum distance between two points, which are clearly seen as separate points, at a standard distance from the observer. Humans with normal acuity can distinguish two points separated by an angle of one minute of arc. This is usually expressed clinically as the reciprocal of the angle or 1.0; values less than one indicate reduced acuity. Some people can distinguish separations as small as 0.5 to 0.33 minute of arc and therefore have acuities of 2.0 to 3.0.Because the retina is an array of receptors and not a continuous sheet, two point sources of light will only be seen as separate if the light from each one falls on a different receptor and the receptors are connected to separate communication lines to the brain, i.e., to separate ganglion cells. From what you have already learned about the retina, you should immediately conclude that acuity is greatest in the fovea. This is illustrated in Figure 7-10, where visual acuity has been plotted against position of a test spot on the retina. Acuity is 1.0 in the fovea and falls to 0.1 within 30 degrees of the fovea. Acuity is, of course, zero in the blind spot where there are no receptors.
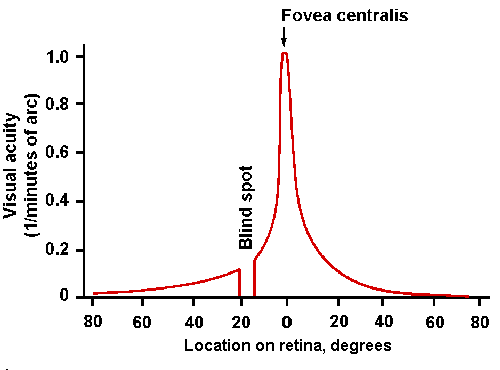 |
Fig. 7-10. Visual acuity as a function of position on the retina. Note position of blind spot (abscissa) and maximum acuity (1.0) at fovea centralis. (Gruesser O-J, Gruesser-Cornehls U: Physiology of vision. In Schmidt RF [ed]:Fundamentals of Sensory Physiology. New York, Springer-Verlag, 1978) |
The fovea is specialized for detail vision in several ways: (1) the cones are more slender and more closely packed than elsewhere in the retina, (2) each cone has its own labeled line in the optic nerve, and (3) the blood vessels and extra cells are pushed out to the side of the fovea, reducing their light scattering to a minimum. These factors combine so that the acuity near the edge of the macula is only half that at the fovea, and the acuity at the periphery of the retina is only one-fortieth that at the fovea. The fovea may also act as a lens to magnify the image in that region. Recall that the fovea is concave. Because the retina has a different (greater) refractive index than the vitreous, it could act as a diverging lens that would magnify the image on the dense array of cones, further increasing acuity in the fovea.
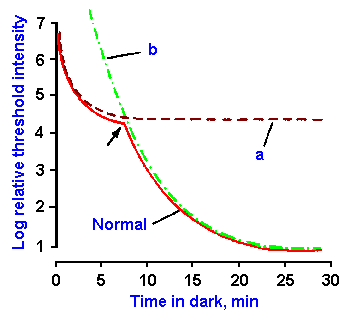 |
Fig. 7-11. The time course of dark adaptation. The threshold in log units for vision is plotted as a function of time in the dark. Adaptation in the normal eye (solid curve) occurs in two phases. When colored light is used, sensation is chromatic to the arrow, achromatic thereafter. Curve a is the adaptation curve for a night-blind person. Curve b is the adaptation curve for a completely color-blind person. (Gruesser O-J, Gruesser-Cornehls U: Physiology of vision. In Schmidt RF [ed]: Fundamentals of Sensory Physiology. New York. Springer- Verlag, 1978) |
Adaptation
Everyone has experienced the changes in visual sensitivity that occur when going from a situation of bright light into one of dim light, such as when entering a darkened theater. At first not much can be seen, but, after a period of time, a considerable sensitivity returns. This increase in sensitivity of the retina is called dark adaptation. The reverse condition, a supersensitivity, followed by a decrease in sensitivity on emerging from the dark also occurs, and the decrease in sensitivity is similarly termed light adaptation. Figure 7-11 shows the time course of dark adaptation (solid curve). In the graph, the log of the threshold for vision is plotted against time in the dark on the abscissa. Notice that the threshold decreases in two phases, one lasting about 7 minutes and the other beginning at about 7 minutes and completed by 30 minutes. During this time, the threshold has decreased by nearly one million-fold. Recall that the light reflex can produce only a 16-fold change. Clearly, the majority of compensation for changes in light intensity is accomplished through adaptation.The two phases probably represent changes first in the cones and then the rods. There are four pieces of evidence for this conclusion: (1) if colored light is used, the sensation is chromatic to the break point (arrow in Fig. 7-11) and then it becomes achromatic, (2) if red light (rods are insensitive to red light) is used and directed only at the fovea, the second phase is absent (Fig. 7-11, curve a), (3) in persons with severe cases of congenital night-blindness (nyctalopia), in whom rod function is apparently completely absent, the second phase is absent (Fig. 7-11, curve a), and (4) individuals lacking cones, i.e., completely color-blind persons, show only the second phase of adaptation, but starting at zero time in the dark (Fig. 7-11, curve b).
Dark adaptation results from the fact that vision is a photochemical process. When light strikes the retina, some of the photons are absorbed by the molecules of the visual pigment embedded in the membranous disks of the receptors. When a molecule absorbs a photon it changes to a higher energy state. In this state, it may undergo a configurational change leading to decomposition of the pigment molecule. The visual pigment of rods, rhodopsin, consists of a protein called opsin and an aldehyde of vitamin A1 called retinene 1. When rhodopsin is decomposed to opsin and vitamin A1, the conductance of the lamellated disk membrane (the membrane that contains the pigment) for calcium is thought to increase, allowing calcium to diffuse into the intracellular space, stabilize the outer receptor membrane and reduce the conductance for small ions, particularly sodium. It is this reduction in the membrane conductance that is responsible for the receptor potential of the rods and cones. Reducing the sodium conductance results in an hyperpolarization of the membrane.
In this decomposed or "bleached" condition, the pigment cannot absorb another photon and, therefore, cannot participate further in the visual process. However, there is a process, that involves active pumping of calcium into the membranous disks, that results in the reassembly of the pigment. The sensitivity of the receptor is a direct function of the amount of unbleached pigment it contains. When moved into the light, pigment is bleached faster than it is reformed and there is a reduction in sensitivity. This is light adaptation. If enough pigment is bleached, a temporary blindness develops. On the other hand, when the light-adapted person goes into the dark, his sensitivity to light is low because most of his pigment is in the bleached condition, but because the pigment is being bleached at a lower rate, the regeneration process can reform the pigment faster than it is being bleached. The result is a gradually increasing quantity of unbleached pigment and thus a gradual increase in sensitivity or dark adaptation. Under steady illumination, an equilibrium condition is reached between bleaching and reformation processes, and sensitivity of the retina stays constant. The exact position of this equilibrium depends upon the level of illumination.
At first, it may seem puzzling that the rod adaptation curve of Figure 7-11 is delayed. One reason for this is that cone pigments regenerate more rapidly than rod pigments. Also, the cone system seems to exert a suppressive control of the rod system, as mentioned earlier. Clinically, this suppressive effect is manifest in the photophobia of the totally color-blind person. The totally color-blind person functions well in dim light but has an abnormally (painfully) high sensitivity in bright light. This presumably reflects the lack of suppression of rods by the cone system.
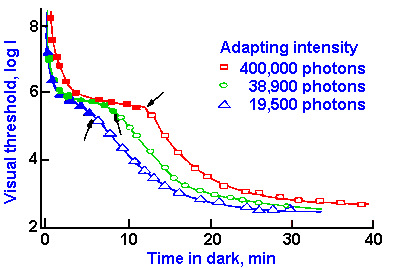 |
Fig. 7-12. The course of dark adaptation as measured with violet light following different degrees of light adaptation. The filled symbols indicate that a violet color was apparent at the threshold, whereas the empty symbols indicate that the sensation at threshold was colorless. (Hecht S, Haig C, Chase AM: J Gen Physiol 20:831-850, 1937) |
The break point in the adaptation curve of normal subjects (arrow in Fig. 7-11) comes at a later time in the adaptation period when a brighter light is used in the period before the dark adaptation is measured. This is illustrated in Figure 7-12, where the same data used in Figure 7-11 are plotted (19,500 photons) along with curves for two higher intensities of light during the pre-dark adaptation period. Again, the break points are indicated by arrows. This delay reflects an increase in light adaptation in brighter light that in turn reflects a greater amount of bleaching of the photosensitive pigments.
It is the cone receptors that contain the visual pigments that are sensitive to the color of light. In the 1800s, two physiologists, Thomas Young and Hermann von Helmholtz, proposed that there are three fundamental color (hue) sensations in human vision, namely red, green, and violet (or blue), and that there are three different kinds of color receptors, one for each of these colors. Excitation of a given kind of receptor leads to the sensation of the appropriate color, and all other color sensations result from simultaneous excitation of more than one of these receptors. Thus, equal excitation of all three receptor types leads to the sensation of white light, whereas the appropriate amounts of excitation of red and green receptors yields the sensation of yellow, and so on. This theory, called the Young-Helmholtz, or the trichromatic theory, is the generally accepted theory today. |
Fig. 7-13. Contributions of the individual color-receptor mechanisms to the total foveal sensitivity of a single human subject. In this particularly blue-sensitive observer, when the total foveal sensitivity is given a maximal height of 1.0, the heights of the B, G, and R curves are 0.053:0.575:0.542. In the average observer, the blue component is only about one-third as high as that shown here. (Wald G: Science 145:1007-1016, 1964) |
In the early 1960s, George Wald (a visual physiologist who later was awarded the Nobel prize) measured the sensitivity of the pigments of the fovea using psychophysical methods. His results are shown in Figure 7-13. Overall spectral sensitivity of the cones was measured by shining lights of various wavelengths on the fovea and determining the threshold strength for each. A plot of these thresholds is seen in the bold (upper) curve in the figure. Then, the fovea was illuminated (adapted) with blue-green light (blue and green), presumably bleaching the green and blue pigments, leaving only the red functioning, and a new spectral sensitivity curve was measured (curve R in the figure). Likewise, the fovea was adapted to purple (blue and red) light and the green pigment curve (curve G) was measured; the fovea was adapted to yellow (red and green) light and the blue pigment curve (curve B) was measured. The blue pigment curve shows peak sensitivity to light of wavelength about 440 nm, the green pigment to light of wavelength about 550 nm, and the red pigment to light of wavelength about 580 nm. It should be noted that only the red pigment is sensitive at all to light of wavelength longer than 650 nm, though there is a rather broad overlap in sensitivity throughout the remainder of the visible spectrum. In addition, the sensitivity of the blue pigment is very much less than that of either of the others.
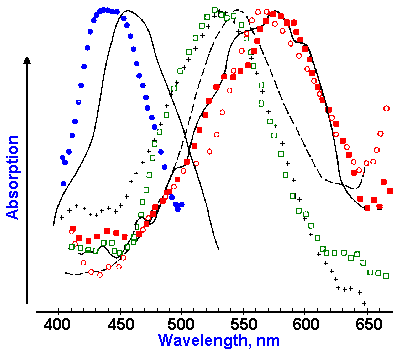 |
Fig. 7-14. Absorption difference spectra for eight primate cones. Two human cone spectra are indicated by the solid curves. Each symbol represents a different cone. (Wald G: Science 145:1007- 1016, 1964) |
Spectral sensitivities have also been measured for individual cones. In this case, the absorption of light of various wavelengths by the pigment in a single cone is the variable measured instead of the sensitivity of the person as indicated by his verbal report, as used by Wald. The absorption of light of a given wavelength is determined by shining a light of that wavelength into the eye and measuring how much comes back out. The difference between that amount and the amount that went in is the amount absorbed by the pigments. Plots of absorption spectra for eight cones, including two from human retinae and six from other primates, are shown in Figure 7-14. There are clearly three different kinds of absorption spectra represented in the figure, with peak absorptions at about 445, 535, and 570 nm. These curves are not bad matches for those in Figure 7-13, considering that they were obtained by different methods and that one represents individual receptor responses, whereas the other represents a pigment population response.
Evidence indicates that the three types of cones are not distributed uniformly across the cone portion of the retina. Blue cone density is greatest at the edge of the fovea (the human fovea is 1 degree in diameter) and falls off markedly by 10 degrees from the center. Red and green cones are distributed most densely in the fovea, with the density of green cones being greater than that of red cones. Over the entire retina the green cones are more dense than the red, which are in turn more dense than the blue. Furthermore, blue cones appear in regular arrays, whereas the others appear to be distributed randomly.
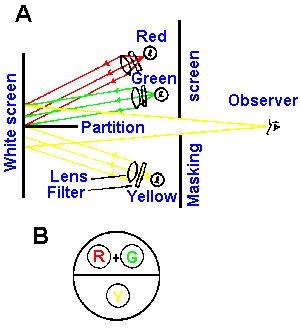 |
Fig. 7-15. A. Representation of a color mixture experiment. The top half of the bipartite field is illuminated by the additive combination of red and green light. The bottom half of the field is illuminated with yellow light. B. View of the field as seen by the observer. |
There are two ways of producing the various colors of the visible spectrum. One way is to shine a light of the appropriate wavelength in the eye. For example, the sensation of yellow light can be produced by shining a light of wavelength 590 nm into the eye. Alternatively, suitable amounts of light of three other appropriate wavelengths can produce the same sensation when shown into the eye at the same time. The same sensation of yellow light can be produced by mixing light of wavelength 545 nm (green) and 670 nm (red). (No blue light is required to produce the sensation of yellow, but for other sensations a certain amount of blue light may be necessary.) That this is true is indicated by the following experiment (Fig. 7-15). The observer looks through a hole in a partition at a white surface that is bisected by a black partition. What he sees is shown in Figure 7-15B. A yellow light of 589.3 nm is shown on the lower half of the field, and the observer reports seeing a yellow light. A light of 546 nm is shown on the upper half, and he reports seeing a green light; this is turned off and a light of 670.8 nm is shown on the upper half which he reports to be red. When both the red and green lights illuminate the upper field at the same time at the appropriate relative intensities, he reports that both the upper and lower fields are the same color, namely yellow. This is the basis of function of the anomaloscope used to detect color blindness.
Red, green, and blue were taken as the primary hues in this example, because these are the ones used by the human eye, but, from a physical point of view, any three other pure spectral hues could also be used. For the human eye, any color experienced can be produced by a simple additive mixture of the correct proportions of red, green, and blue light.
Color blindness
Color blindness can be partially understood in terms of the mechanisms of color vision discussed above. Essentially, there are three types of individuals with respect to color vision, as follows:Types of Color Vision
- Trichromats
- Normal
- Protanomalous
- Deuteranomalous
- Dichromats
- Protanopes
- Deuteranopes
- Tritanopes
- Monochromats
- Totally color blind
There are persons, including those with normal color vision, who have three visual pigments that they use in their color perception. These are the trichromats. Those in a second class, the dichromats, match all the colors they see using only two colors; those in the third class, monochromats, see everything as a shade of gray, because they only see one color, white.
The protanomalous person has all three color pigments, but he has a deficiency in his red responsiveness. In the anomaloscope, he requires at least 1.5 times more red intensity than normal in the upper field to match a yellow in the lower field. The deuteranomalous person has reduced green sensitivity and requires at least four times more intense green than normal to match yellow. This is in fact how these abnormalities in color vision are detected, by measuring the amounts, relative to the normal, of red and green that are required to match a standard yellow. As a consequence of this reduced sensitivity, a person with a color anomaly can identify fewer hues than a normal person can. A normal person can distinguish about 150 different hues, whereas a protanomalous or deuteranomalous person distinguishes only 5 to 25 hues depending upon the severity of the deficiency. In addition, there are some interesting contrast phenomena that show up in anomalous individuals. For example, if a yellow light is shown in the lower field of an anomaloscope, the protanomalous individual identifies it as yellow, but if a green light is simultaneously shown in the upper field, he will report that the lower field is red. This does not happen in normal vision; the lower field remains yellow.
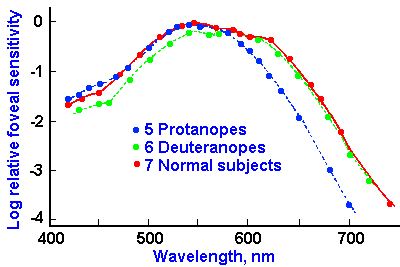 |
Fig. 7-16. Luminosity curves of color-blind and normal subjects. Downward in the graph represents loss of sensitivity, the log of the reciprocal of threshold energy. (Wald G: Science 145:1007-1016, 1964) |
The dichromats present a different and more complicated picture. Figure 7-16 shows the luminosity curves for normal subjects, protanopes and deuteranopes. The protanope lacks red-sensitive pigment altogether. Notice the more rapid fall of the curve for protanopes at the long end of the spectrum (reds). This is the characteristic that distinguishes protanopes; they are completely insensitive to light beyond about 680 nm. The result is that they see little or no red. They have been known to show up at a black-tie affair with a red tie on. The sensation of black is due to the absence of light and is therefore indistinguishable from red (of wavelength greater than 680 nm) by the protanope.
The deuteranopes, on the other hand, are insensitive to green light. There are two types of deuteranopes. There are those who have a reduced sensitivity to light in the middle and shorter wavelengths, which indicates an absence of green pigment (Fig. 7-16), and there are those with normal spectral sensitivity whose problem seems to be that the red and green mechanisms are coupled together to form a single sensory system. This may occur in the retina at the level of the bipolar or ganglion cells, or it may occur more centrally in the visual system. The tritanope is a rare individual who lacks sensitivity to the short end of the spectrum. Some people are of the opinion that tritanopia is not a retinal problem at all, but a disease of the optic nerve. Not enough information is available to say with confidence.
No matter what his deficiency, the dichromat matches all colors he sees with mixtures of only two pure spectral colors. The number of colors remaining chromatic varies with the type of dichromatism, but all of them are interpreted as various saturations of blue and yellow for the protanope and deuteranope, and red and green for the tritanope. For each individual, there is a point of the visible spectrum between his two basic sensations that he matches with what a normal person sees as white. For the protanope this occurs at about 495 nm; for the deuteranope it occurs at 500 nm; for the tritanope it occurs at 572 nm. One unanswered question is why the protanope (and by analogy, the deuteranope) sees yellow at the middle (long end) of the spectrum and not green (red). It is possible that the pigment present (either red or green) excites both red and green mechanisms in the central visual system resulting in a sensation of yellow.
Kalmus (1965) has reported an interesting individual who was a dichromat in one eye and trichromat in the other.
|
The cause of anomalous trichromacy (protanomaly or deuteranomaly) is usually stated to be an abnormally small amount of the appropriate pigment in the cones. This cannot be the case as evidenced by the following experiment:
A normal observer adjusts red and green intensities to match a yellow in an anomaloscope, as described above. He then looks away from the anomaloscope into an intense red light, bleaching the red pigment. If he looks immediately back into the anomaloscope, he will see green light in both the upper and lower fields, and both are the same green without adjustments of the intensity. If he adapts to green light instead, he will see the same red in both fields. This indicates that the anomaloscope cannot distinguish the color-adapted eye from the non-color-adapted eye. We know that color adaptation results from a simple reduction in non-bleached pigment. It follows that if the anomaly were due to such a simple reduction, the anomaloscope wouldn't detect it, but it does.
What then, is the cause of anomalous trichromacy? A protanope can exactly match green with any red of wavelength less than 680 nm simply by adjusting the intensity of the red. As a result, an experimenter can exchange the matching red with the previously presented green, and the protanope will be none-the-wiser. The reason for this is that his green-cone pigment catches the light equally and experiences no change when they are switched. The same trick cannot be played on a protanomalous individual; he sees the switch. His green pigment cannot detect it; this suggests he may have some anomalous pigment. In fact, such anomalous pigments have been identified by using the trick above to measure their sensitivity. The anomalies are probably due to a malfunction in the process for making the pigment. The correct red is replaced by an anomalous one that is less sensitive in the red region in the protanomalous individual; and, in the deuteranomalous individual, the green is replaced by anomalous pigment that is more like the red pigment. The sum of these effects is that the remaining normal pigment (green in the case of protanomaly) and the anomalous pigment sensitivities are much closer together than the two normal pigments (red and green), and the individual has a harder time distinguishing colors in this region of the spectrum. He thus requires more intensity of red to excite the anomalous pigment than he would to excite a normal red pigment to achieve the proper balance of green and red (anomalous red pigment) excitation.
Protan and deuteran disturbances (terms for protanomaly and protanopia and for deuteranomaly and deuteranopia, respectively) are simple, sex-linked recessive traits. The trait is linked to the X chromosome, and so the condition is most common in males. Females show the condition only if they have it present on both X chromosomes. These conditions occur in 1-13% of males, depending upon the country (USA about 8%), but only 0.5% females. The normal condition is dominant to the protan or deuteran, and the anomalous condition appears to be dominant to the anopic. The control of green and red pigments seems to be at two separate loci on the chromosome, as indicated by family history data and the existence of women with normal color vision, but with one deuteranope and one protanope son. In the latter case, the women must have carried one defective gene of each type, yet neither was expressed in their phenotype. Deuteranomaly is the most frequently encountered defect, with protanomaly, deuteranopia, and protanopia less but about equally frequent. Tritanopia only occurs in about 0.002% of the male population, and thus it is a rare event.
The last color vision defect is monochromatism. This usually shows up in the form of total color blindness in which all colors are perceived as shades of gray. People with this problem are photophobic, as we have mentioned, and show only scotopic luminosity curves. There are cases of subtotal monochromatism in which blue cones are not or only minimally involved or in which mainly macular cones are affected. The result is nearly the same because of the similarity of the blue cone and rod spectral sensitivities and because the macula is the major region used in vision in humans.
| Physiology of single visual neurons |
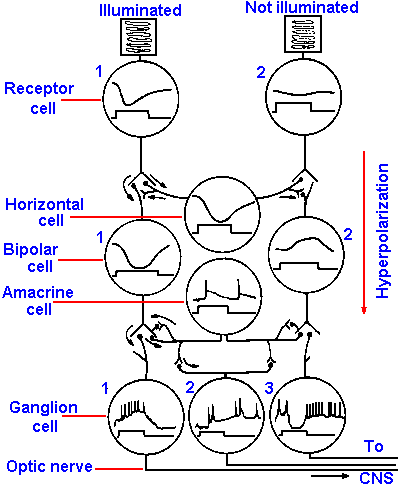 |
Fig. 7-17. Synaptic organization of the vertebrate retina and responses of retinal neurons. The receptor on the left is illuminated, that on the right is not. The intracellularly recorded response from each cell is illustrated in the circle corresponding to its cell body (upper trace) along with a monitor of when the illumination was present over the left receptor. The responses were recorded in the retina of Necturus. (Dowling JE, Werblin FS: Vision Res Suppl 3:1-15, 1971) |
Visual neurons in the retina
The receptor cells and the bipolar cells of the retina respond to light with graded, electrotonic responses rather than all-or-nothing action potentials. The graded responses in the receptors are the result of the photochemical process, but those in the bipolar cells are synaptically driven. Furthermore, it may be surprising that the receptors respond to light with an hyperpolarizing receptor potential, that is accompanied by an increase in membrane resistance. Figure 7-17 shows a schematic diagram of a section of the retina with two receptors, one illuminated, the other unilluminated. The responses of the receptors, bipolar, ganglion, horizontal and amacrine cells are shown in circles representing each cell type. The time when a small spot of light was turned on is indicated by the upward deflection of the lower trace of each pair and the response of the cell by the upper trace. The hyperpolarizing responses of the illuminated receptor and its subjacent bipolar cell are illustrated as are the action potentials generated by the ganglion cell. It is now known that in the dark there is a constant inward Na+ current (dark current) flowing through the outer segment membrane and an outward current near the junction with the bipolar cell. This keeps the cell partly hypopolarized, and transmitter substance is continuously released onto the bipolar cell hypopolarizing it. The light flash decreases the dark current by the action of calcium to reduce membrane conductance, hyperpolarizes the cell relative to its dark state and decreases the amount of transmitter released onto the bipolar cell. If this scenario is correct, then adding excess magnesium to the bathing solution should cause bipolar and ganglion cells to behave as if their receptors were illuminated because excess magnesium blocks the release of transmitter substances at chemical synapses. A release of transmitter substance blocked by magnesium should be the same as one blocked by light, and it is. This scheme accounts mechanistically for the curious hyperpolarizing receptor potentials. But why are the receptor potentials hyperpolarizing rather than hypopolarizing as in other types of receptors? It could be that we have identified the wrong stimulus! Perhaps it is darkness, not light, that is the stimulus. During evolution, when these receptors developed, the animals were living in the oceans and there were few land plants to cast shadows. Therefore, shadows cast by predators were relatively more frequent and predicted disaster. If an animal could detect these shadows and respond quickly (by swimming away), he would have a survival advantage. Such a significant advantage would survive the evolutionary process. So . . . the receptors were probably initially shadow-detectors, and the proper stimulus is darkness. In that case, the receptors hypopolarize in response to the dark stimulus!The major line of transmission of information is from receptor to bipolar cell to ganglion cell and then to the brain, but the amacrine and horizontal cells provide lateral transmission lines that can produce the complicated center-surround receptive fields of ganglion cells. The horizontal cells receive their inputs from receptors as do the bipolar cells and, like the bipolar cells, they generate no spikes. Horizontal cell outputs inhibit (reduce transmission at) nearby unilluminated receptor-bipolar cell synaptic junctions. It has been suggested that they may enhance contrast by strongly turning off unstimulated bipolar cells. Amacrine cells produce action potentials and enter into reciprocal synaptic relations with bipolar cells.
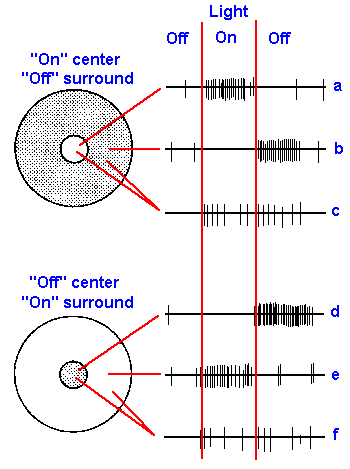 |
Fig. 7-18. Receptive fields of two retinal ganglion cells. Fields are circuluar areas of the retina surrounded by an annulus of different properties. The cell in the upper part of the figure responds when the center is illuminated (on-center, a) and when the surround is darkened (off surround, b). The cell in the lower part of the figure responds when the center is darkened (off-center, d) and when the surround is illuminated (on-surround, e). Both cells give on- and off- responses when both center and surround are illuminated (c and f), but neither response is as strong as when only center or surround is illuminated. (Hubel DH: Sci Amer 209:54-62, 1963) |
The response recorded from ganglion cells following light stimulation of the receptors is more conventional. Hypopolarizing synaptic potentials initiate trains of action potentials that propagate along the ganglion cell's axon. Ganglion cell responses are of three types: (1) they respond (either discharge or increase their rate of discharge) only when the light is turned on: (2) they respond only when the light is turned off; or (3) they discharge only at the beginning and end of a light period. These are called on-responses, off-responses and on-off-responses, respectively. The receptive fields of ganglion cells are usually circular areas of retina, as shown in Figure 7-18. Illumination within the small central area produces an on-response in some cells (a), an off-response in others (d). In a, the neuron responds when the light goes on, but gives no response when it goes off (on-center); in d, another neuron does not respond when the light goes on, but does respond when it goes off (off-center). Illumination of an annular, fringe area produces an off-response in the first type of ganglion cell (trace b, off-surround) and an on-response in the second (trace e, on-surround). When a ganglion cell fails to discharge at an on- or off-transient of the light, it is not because it is not being excited, but because it is being inhibited, as is indicated by the fact that illumination of both center and surround simultaneously produces both a reduced on-response and a reduced off-response (c and e) compared to those produced by illuminating either center or surround alone. This effect is termed lateral inhibition or surround inhibition.
A look back at Figure 7-17 may help to understand how responses of ganglion cells are generated. The illuminated receptor cell 1 hyperpolarizes, causing synaptic transmission to be reduced to bipolar cell 1 and the horizontal cell. Transmission from bipolar cell 1 to ganglion cell 1 causes an hypopolarization that occurs when the light comes on (on-responses). Transmitter substance is released by bipolar cell terminals in response to hyperpolarization; this is opposite to the situation at most chemical synapses (see Chapter 13). Receptor cell 1 is in the center of the receptive field of ganglion cell 1, but it is in the surround of ganglion cell 3. The hyperpolarization of the horizontal cell by illumination of receptor cell 1 causes bipolar cell 2 to hypopolarize. This hypopolarization prevents the release of transmitter substance from bipolar cell 2 onto ganglion cell 3, and the ganglion cell does not discharge. Actually, the ganglion cell is also inhibited (prevented from firing by hyperpolarization), perhaps through the amacrine cell. When the illumination of receptor cell 1 is turned off, bipolar cell 2 repolarizes. This repolarization is equivalent as far as the transmitter release mechanism is concerned, to an hyperpolarization of the bipolar cell; it causes the release of transmitter substance onto the ganglion cell, which discharges (as the light goes off). This mechanism would account for on-center cells; presumably the mechanism for off-center cells would be similar.
Both rods and cones release the transmitter substance L-glutamate at their terminals on bipolar cells. They do this in response to hyperpolarization when they are illuminated. The L-glutamate produces an hyperpolarization in "off" bipolar cells and an hypopolarization in "on" bipolar cells. Many people think that a given transmitter substance always produces the same effect in postsynaptic cells (for example, acetylcholine is thought [by some people] always to be excitatory and GABA always inhibitory). In the receptor-bipolar cell synaptic arrangement, we see a clear indication that this idea is incorrect. The L-glutamate activates a different kind of receptor in "off" and "on" bipolar cells. In "off" bipolar cells, activation of the KA/AMPA receptor by L-glutamate produces hyperpolarization. In "on" bipolar cells, the L-glutamate activates L-AP4 receptors to produce hypopolarization.
The first stage of processing of color information is apparently at the level of the ganglion cell, at least this is the first one studied to date. Color-sensitive ganglion cells are sometimes called color opponent cells, because cone mechanisms of different spectral sensitivities (i.e., red, green, and blue) interact in them antagonistically. Most of these cells have concentric receptive fields like other ganglion cells, with either on- or off-centers, and the centers are sensitive to red, blue, or green. The annular surround of an on-center gives an off-response for one of the other cone pigments. Cone mechanisms neighboring each other in the spectrum appear to oppose each other so that red centers have only green surrounds and blue centers have only green surrounds, but green centers can have either red or blue surrounds. Red-blue interactions have not been found. As an example, consider a red on-center, green off-surround cell. This cell discharges when the center of the receptive field is illuminated with red light; it is inhibited when the center is illuminated with green light, but discharges when a green light is extinguished after shining on the periphery; and it is inhibited by illuminating the periphery with red light. The result of these interactions is a rather narrow spectral response of the central portion of the receptive field compared with the absorption curve for the pigments. Thus, some of the very broad overlap in the absorption spectra of the neighboring pigments is eliminated by the nervous system, which forms six fairly narrow spectral channels of information with both on and off information about the image.
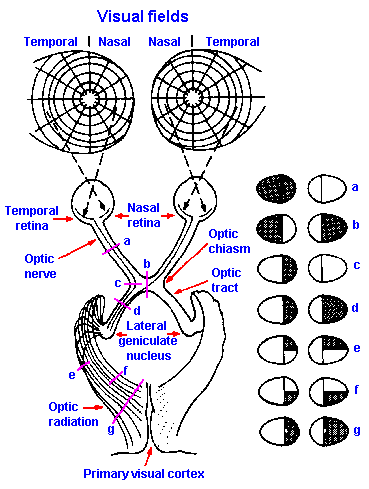 |
Fig. 7-19. The anatomic organization of the visual pathway from the retina to the visual cortex. Lesions of the visual pathway (a-g) produce defects in the visual fields as indicated at the right. (Homans A: Textbook of Surgery, 5th ed. Springfield, IL. C.C. Thomas, 1941) |
Visual neurons outside the retina
The axons of the ganglion cells form the optic nerves, which, after leaving the eyeball, proceed toward the brain until they come to the optic chiasm, where the optic nerves divide. Fibers from the nasal half of the retina cross to the opposite side of the brain; fibers from the temporal half go to the same side of the brain (Fig. 7-19). Past the chiasm, crossed fibers from the contralateral eye join the uncrossed fibers from the ipsilateral eye to form the optic tract. Fibers in the optic tract, which are still the axons of retinal ganglion cells, then proceed to the thalamus, where they end on cells of the lateral geniculate nucleus. The fibers of this nucleus project to neurons of the calcarine area of the occipital cortex. This is area 17, as enumerated by the anatomist, Brodmann, and is often referred to as striate cortex because of its characteristic striped appearance under the microscope. There are also, direct projections to adjacent areas 18 and 19, often referred to as visual association areas.The receptive fields on the retina of single cells in the lateral geniculate nucleus are similar to those of ganglion cells. Retinal receptive fields of geniculate cells have excitatory centers and inhibitory surrounds or vice versa, but the fields are larger, probably due to convergence of more than one ganglion cell on each geniculate neuron. Despite the presence of both crossed and uncrossed tract fibers in the geniculate, each cell there has a receptive field associated with only one eye.
With respect to spectral sensitivity, geniculate neurons can be put into one of two categories: (1) narrow-band cells and (2) broad-band cells. Narrow-band cells respond with an increase in their discharge rate when their receptive fields are illuminated with light from part of the visible spectrum, and they decrease their discharge rate when their fields are illuminated with light from other parts of the spectrum. The majority of narrow-band cells either increase their response rates when their receptive fields are illuminated with red light and decrease their rates when illuminated with green light, or they increase their rates when illuminated with green light and decrease them when illuminated with red light. The remainder of the cells increase their discharge rates when illuminated with yellow light and decrease them when illuminated with blue light or increase their rates for blue light and decrease them for yellow light. No red-blue cells have been found. In most respects, the narrow-band cells are not very different from color-opponent ganglion cells except for the size of their receptive fields and the width of the spectrum to which they respond. The peak sensitivity in terms of wavelength of the excitatory and inhibitory responses of geniculate cells corresponds nicely to the peak absorption spectra of the three cone pigments. Broad-band cells receive a uniform excitation or inhibition when their receptive fields are illuminated by light of all wavelengths and white light.
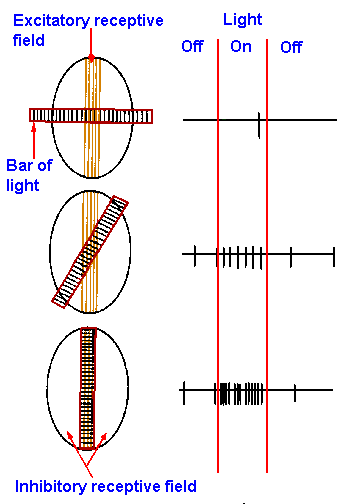 |
Fig. 7-20. The receptive field on the retina (ellipse) of a simple cell in the visual cortex is shown at the left with a bar of light superimposed on it at various angles. The vertically striped portion of the receptive field is the excitatory area (excitatory receptive field), illumination of which excites the cell; the unhatched portion is the inhibitory area (inhibitory receptive field), illumination of which inhibits the cell. The bar of light is striped across its length. The responses of the cell to the three different orientations of the light are shown at the right. (Hubel DH: Sci Amer 209:54-62, 1963) |
The retinal receptive fields of neurons in the
occipital cortex are complicated. Some visual cortical
neurons (non-oriented neurons) possess receptive fields
not particularly different from those of geniculate
neurons; they have circular receptive fields and respond
equally to stimuli of all orientations. However, the
receptive fields of most cortical neurons are not circular,
most are arranged as parallel barlike excitatory and
inhibitory areas with straight, rather than circular
borders. Cortical cells with this characteristic receptive
field are termed simple cells. In one sample of cortical
neurons, two-thirds of the cells had simple receptive
fields. These cells can have two inhibitory regions
flanking an excitatory region, as in Figure 7-20; the
reverse situation can occur; or there can be just two
regions side by side, one excitatory, the other inhibitory.
As a result the cells respond best to narrow bars of light
oriented in a particular direction across the retina.
Sample recordings are shown for three different
orientations of such a bar in Figure 7-20. Rotation of
the bar has two effects: (1) it reduces the excitation,
because less of the excitatory area is illuminated and (2)
it increases the inhibition, because more of the inhibitory
area is illuminated. The result is that the cortical neuron
responds less well when bars are not in their preferred
orientation.
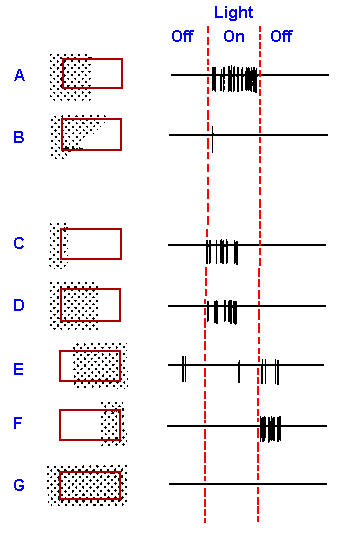 |
Fig. 7-21. The response of a "complex" cortical cell in the visual area to various illuminations of its large receptive field (8 x 16 degrees) indicated by the rectangle. The stippled area is the area illuminated, and the response of the cell to the illumination and its cessation are shown on the right. (Hubel DH, Wiesel, TN: J Physiol (Lond) 160:106-154, 1962) |
In general, complex cells seem to be responsive to lines of sharp contrast. However, unlike simple cells that show only on and off-responses, complex cells respond in a sustained manner to moving lines. They respond better to a line moving in one direction than to the same line moving in the opposite direction across their receptive fields. Figure 7-22 shows the 2-sec records of the responses (on the right) of a complex cell to a bar of light (stippled rectangle) moving at various orientations with respect to the receptive field (3/8° x 3/8°, indicated by the broken square), as shown on the left. The responses of the cell to movement, first in one direction and then the other (arrows), are indicated at the right. For this cell, the best response occurs when the long axis of the bar is parallel to the long axis of the receptive field and the bar is moving toward the right. Speed of movement of the bar also seems to be an important parameter for some cells.
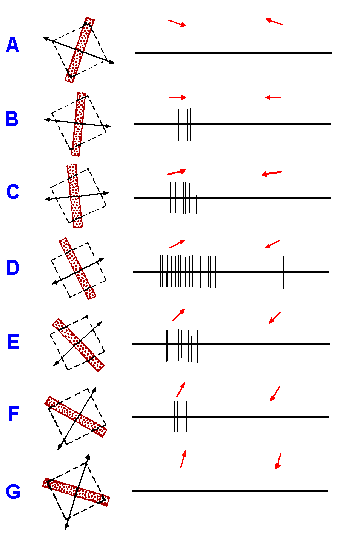 |
Fig. 7-22. Responses of a complex cell in right striate cortex (layer IVA) of Macaque to various orientations of a moving black bar. Receptive field in left eye is indicated by interrupted rectangles, approximately 3/8 x 3/8 degree in size. Duration of each record, 2 sec. Arrows indicate direction of stimulus motion. (Hubel DH, Wiesel TN: J Physiol (Lond) 195:215-243, 1968) |
A third class of visual cortical cells, the hypercomplex
cell, is much like the complex cell in its response to lines of
light or the edges of bright objects, but with the added
complexity that it turns off if the line or edge is too long.
This is clearly seen by reference to Figure 7-23 which shows
responses of a cell whose receptive field (2° x 2°, indicated
by the broken square) was illuminated with a 1/8°-wide bar of
light moving at 3°/sec. Up to a certain point, increasing the
length of the line increases the response, but beyond that
point, increasing the length decreases the response. In short,
they behave as if the excitatory receptive field is bounded on
the ends by an inhibitory field, but no effects (either
excitatory or inhibitory) on cell discharge are seen by
stimulating these boundary regions alone. In the case of
Figure 7-23, the right
inhibitory flank is weaker
than the left one. Many
recent publications have
dropped the term
hypercomplex cells, and have substituted they term "complex
cells with end-inhibition." The cells are still the same as before.
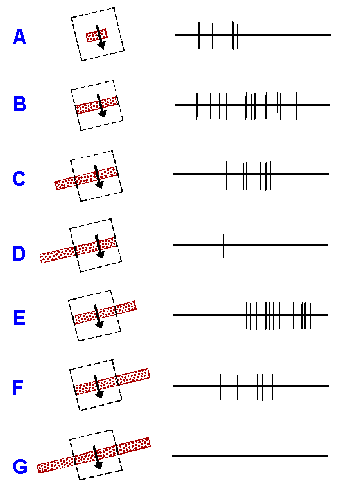 |
Fig. 7-23. Hypercomplex cortical neurons (area 19 of cat). Stimulation of ipsilateral eye. Excitatory portion of retinal receptive field roughly 2 x 2 degreees, represented by interrupted rectangle. (A-G) response (right) to slits 1/8 degree wide and of various lengths moved downward across receptive field at about 3 degrees/sec (left). Note that inhibitory flank on right of excitatory region is weaker than that on left. Sweep duration, 1 sec. (Hubel DH, Wiesel, TN: J Neurophysiol 28:229-289, 1965) |
The color sensitivity of visual cortical neurons is much the same as that of geniculate neurons. There are some with broad-band sensitivity and some with narrow-band sensitivity. Most of the narrow-band cells (those presumably signaling color information) are either non-oriented or simple cells. They are excited by light of some wavelengths of the visible spectrum and inhibited by some other wavelengths, much as described earlier for geniculate cells. Other cortical neurons, mainly some complex cells, are excited by illuminating their receptive fields with light from narrow bands at one end or the other of the spectrum. These narrow-band cells make up about 54% of all those cells in samples from the foveal representation in the striate (visual) cortex. The broad-band cells are very similar to broad-band cells in the lateral geniculate nucleus.
We have as yet no idea how color is perceived, but its perception seems to involve contrasting excitation and inhibition from the different types of cones or rather cone systems. There is some indication that as a cell's color specificity increases, its spatial selectivity decreases, implying that there are separate mechanisms (cells) for color and form perception.
| Clinical aspects of visual function |
It should be noted, by reference to Figure 7-19, that light striking the nasal retina comes from the temporal visual field(7) and that striking the temporal retina comes from the nasal visual field. The monocular visual field is the total extent of the visual world that is seen by one eye when it is not moving. It is customary to describe the defects of vision in terms of losses in the visual field, the measurement of which is called perimetry. In the maps of Figure 7-19, the basic reference is a line perpendicular to a tangent to the center of the surface of the cornea and passing through the center of the fovea. This is the visual axis. The visual axis is marked in the figure as the point in the center of the concentric circles. The vertical diameter of the circles is the same for both eyes so that there is considerable overlap in the visual fields of the two eyes, the basis for stereoscopic vision.
At the right of Figure 7-19 are shown defects of the visual field that would develop from lesions at the levels indicated by the corresponding letters on the diagram to the left. It is possible to work out these defects from knowledge of the anatomy of the visual pathway. Just keep in mind the following points: (1) the retinal image is inverted top to bottom and left to right with respect to the visual field, (2) fibers from the nasal half of the retina cross over in the optic chiasm, (3) fibers from the upper part of the retina segregate from those from the lower, (4) upper retinal fibers from one eye stay with the upper retinal fibers from the other eye; the same is true for the lower retinal fibers, (5) fibers from the upper part of the retinae are found dorsally in the superior optic radiation passing through the parietal lobe, those from the lower part are ventrally in the inferior radiation (Meyer's loop) passing through the temporal lobe, and (6) the macula may be represented on both sides of the cortex.
Loss of half of the visual field is called hemianopsia. If the visual field is half-lost on either the right or the left, as for example in left optic tract section, Figure 7-19d, the loss is called homonymous hemianopsia, in this case, right homonymous hemianopsia. If the loss is in both temporal fields, it is called bitemporal hemianopsia (Fig. 7-19b). Likewise, the nasal counterpart in binasal hemianopsia. A quadrantanopsia results from interruption of either the superior or inferior optic radiation (clinically this usually involves a lesion in either parietal or temporal cortex) or alternatively from a partial lesion of the calcarine cortex, lower quadrants in the lingual gyrus, upper in the cuneate gyrus (Fig. 7-19e,f). The representation of the macula on both sides would result in macular sparing after a complete lesion of one visual cortex. In this condition, vision remains in the entire macula, as illustrated by the peculiar picture in Figure 7-19g. There are some investigators who believe that macular sparing is an artifact of inadequate control of eye movements during perimetry. As yet, this argument is unsettled.
For a long time, it has been generally agreed that complete removal of the visual cortex in man leads to total, permanent blindness in the contralateral visual field, i.e., contralateral to the removed cortex. In the last ten years, it has become clear that this is not the case in the monkey. With total striate cortex removal, the monkey is still capable of locating briefly presented stimuli in space and can still discriminate visual patterns. With less than total removal, the impairment is best characterized as a reduction in, not a loss of, sensitivity in part of the contralateral field, and, with training, the sensitivity gradually improves. When the lesion involves more than the striate cortex, especially when it includes the posterior parietal and temporal cortices, the animal is reduced to being able to tell only whether the light in its environment is on or off.
In man, there is seldom a pure striate cortex lesion because the tissue is buried in the calcarine tissue. However, in a few cases of pure striate lesions, forced-choice discrimination methods have uncovered considerable visual function. The methods involve instructing the patient to guess the identity or point to the location of a stimulus even though he does not see it. Remaining capacities are much like those in monkeys with similar lesions and include the ability to locate stimuli in space, to differentiate the orientation of lines (vertical versus horizontal versus diagonal, including white on black and black on white), to discriminate shapes (X versus O), and discriminate colors (red versus green). The patients require moderately large stimuli; their performance falls off if the stimuli are too small. For the more difficult discriminations (where the stimuli are more alike, e.g., vertical versus diagonal lines), a slightly longer duration of exposure may be required.
Even though the human with the striate cortex lesion can perform without errors on such discriminations, he still reports that he saw nothing at all and expresses surprise when he is told of his performance. These observations emphasize the fact that information about stimuli goes to more than one center in the nervous system, and they show that what are sometimes called "subcortical relays" really do have functions. A person does not have to be able to report verbally the existence of a stimulus to respond appropriately to it. This sort of observation has led some investigators to parcel visual functions as (1) recognizing, identifying, or examining functions, presumably the territory of cortical visual tissues, and (2) detecting or orienting functions, presumably the territory of subcortical visual tissues. Only time will tell if this sort of distinction has meaning, but it does seem to fit the observations described.
There has been a flurry of activity designed to generate a prosthetic device for blindness. Such devices have been of two sorts: The first represents an attempt to use some other modality with pattern recognition capabilities, and the second, more daring effort, actually attempts to produce a visual image. An example of the first type of device is a television camera whose output is transduced into an array of tactile stimuli that is applied to the skin of the back. The camera is worn on the head, and the picture it sees is perceived by the wearer in sufficient detail that he can avoid obstacles while walking around. On the other hand, it can hardly be claimed that the person has vision.
The second type of device uses an array of photocells or a television camera whose output is applied to an array of small electrodes inserted into the retina or into the visual cortex itself. From what we know of the physiology and anatomy of the retina, it should come as no surprise that stimulation there has not reproduced visual experience. After all, the retina consists of about 106 million receptors, each independently sensitive to light. It seems unlikely that the activity of those receptors could be matched by a small array of electrical stimuli.
Electrical stimuli applied to the visual cortex typically produce sensations described as spots or balls of light that are either stationary or in motion. In no case has the sensation been one of perceiving an object, even when an array of stimulating electrodes was used. Although all these devices have proven useful in helping blind people get around, none of them reproduces the lost vision.
| Summary |
The eye consists of a group of photoreceptors and the optical devices that bring light from outside to a focus on them and regulate the amount of light that strikes them. Refraction is the bending of light in such a way as to bring it to a focus on the retina (in the case of the eye). Most refraction is done by the cornea, but fine adjustments for viewing near objects (accommodation) are accomplished by the lens. The shape of the lens and thus its refractive power is changed by varying the amount of tension in the ligaments that hold it in position. The muscles that reduce this tension are under autonomic control. Hyperopia and myopia are conditions variant from normal (emmetropia) in which the image is focused behind and in front of the retina, respectively. They are corrected by use of either a convex or a concave lens. Astigmatism results from asymmetry in cornea shape. The functions of the pupil are to partially correct for spherical and chromatic aberrations, to adjust depth of field, and to rapidly regulate the amount of light entering the eye over a narrow range of intensities.
The range of wavelengths sensed by the human eye differs in high and low illumination, yielding photopic and scotopic visibility curves. The scotopic curve results mainly from rod sensitivity, whereas the photopic curve results mainly from cone sensitivity. The retina is more sensitive to light on the periphery where the rods predominate. Acuity is greatest in the fovea where cones are packed tightly together; where supporting tissues have been pushed aside; and where the curved surface of the retina may produce some magnification. When a person moves from a light environment to a dark environment, the sensitivity of the eye increases over a period of minutes. This is dark-adaptation and is due to changes in the amount of unbleached pigment in the receptors. Light-adaptation is the reverse process, a decrease in sensitivity with time in the light, and occurs because pigment is bleached faster than it is reformed. There are three types of cones that give color sensitivity: red, green and blue. They have different spectral sensitivities and different distributions across the retina. All color sensations are produced by an appropriate mixture of excitation of the three types of cones. The various forms of color-blindness result from the absence of one type of cone or cone pigment, from pigment anomalies, or from faulty neural connections somewhere beyond the receptors.
Receptor and bipolar cells encode visual information in terms of electrotonic potentials. Ganglion cells have excitatory receptive fields on the retina that are circular, surrounded by an annulus. They give on-, off- or on-off responses to light when illuminated in the circular region and one of the other responses when illuminated in the annulus or "surround." Color is probably coded at this level by color opponent cells. Lateral geniculate receptive fields are like ganglion cell fields but larger. Cortical neurons can be classified as simple, complex, or hypercomplex cells depending upon whether they have stereotyped relationships between excitatory and inhibitory receptive fields, whether they respond differentially to moving lights, and whether the length of the edge of light is critical to discharge frequency.
Even in the absence of visual cerebral cortex there is still some visual function in the contralateral visual field, though it is hardly normal. Stimuli can be located in space; some pattern discrimination and color discrimination are possible. The patient, however, reports seeing nothing.
| Suggested Reading: |
- Baylor DA, Fuortes MGF: Electrical responses of single cones in the retina of the turtle. J Physiol (Lond) 207: 77-92, 1970.
- Dowling JE, Werblin FS: Synaptic organization of the vertebrate retina. Vision Res (Suppl) 3: 1-15, 1971.
- Kaiser PK, Boynton RM: Human Color Vision, 2nd Ed., Washington, D.C., Optical Society of America, 1996.
- Kalmus H: Diagnosis and Genetics of Defective Colour Vision. London, Oxford Univ Press, 1965.
- Lennie P, D'Zmura M: Mechanisms of color vision. CRC Crit Rev Neurobiol 3:333-400, 1988.
- Makous W, Boothe R: Cones block signals from rods. Vision Res 14: 285-294, 1974.
- Neitz J, Neitz M: Color vision defects. In: Wright AF, Jay B (Eds.), Molecular Genetics of Inherited Eye Disorders, Chur, Switzerland, Harwood Academic, pp. 218-257, 1994.
- Osterberg G: Topography of the layer of rods and cones in the human retina. Acta Ophthalmol (Suppl) (Copenhagen) 61: 1-102, 1935.
- Records RE [ed]: Physiology of the Human Eye and Visual System. Hagerstown, MD, Harper and Row, 1979.
- Tessier-Lavigne M: Visual processing by the retina. In: Kandel ER, Schwartz JH, Jessell TM (Eds.), Principles of Neural Science, 4th Ed., New York, McGraw-Hill, pp. 507-522, 2000.
- Weiskrantz L: Some aspects of visual capacity in monkeys and man following striate cortex lesions. Arch Ital Biol 116: 318-323, 1978.
- Wurtz RH, Kandel ER: Central visual pathways. In: Kandel ER, Schwartz JH, Jessell TM (Eds.), Principles of Neural Science, 4th Ed., New York, McGraw-Hill, pp. 523-547, 2000.
Footnotes:
1. Actually, light is an exceedingly complicated and interesting phenomenon. Even casual reading of the popular book, QED, The Strange Theory of Light and Matter, by Richard Feynman (Princeton University Press, Princeton, NJ, 1985) will convince the reader that Newton only scratched the surface.
2. It is unfortunate that the same term is used for the process of forming a focused image and for changes in critical firing level that occur with small prolonged hypopolarizations. The only way to distinguish these usages is by the context in which they occur.
3. There are other ways of accomplishing accommodation. The horse, for example, does it with an irregularly shaped lens that is simply rotated to change the distance from lens to retina..
4. Not everyone agrees that the superior colliculus is involved. There are those who claim it is the pretectal nucleus, which of course is nearby. To be consistent with our neuroanatomists, I have adopted the superior colliculus as a point in this pathway.
5. This is why the sky appears to be blue!
6. The maximum pupil size is about 7 mm in adults and up to 9 mm in children.
7. Here "visual field" refers to the part of the visual world from which light comes to strike that part of the retina.
[TOC] [Chapter 8] [Glossary] [Index] [Abbreviations]